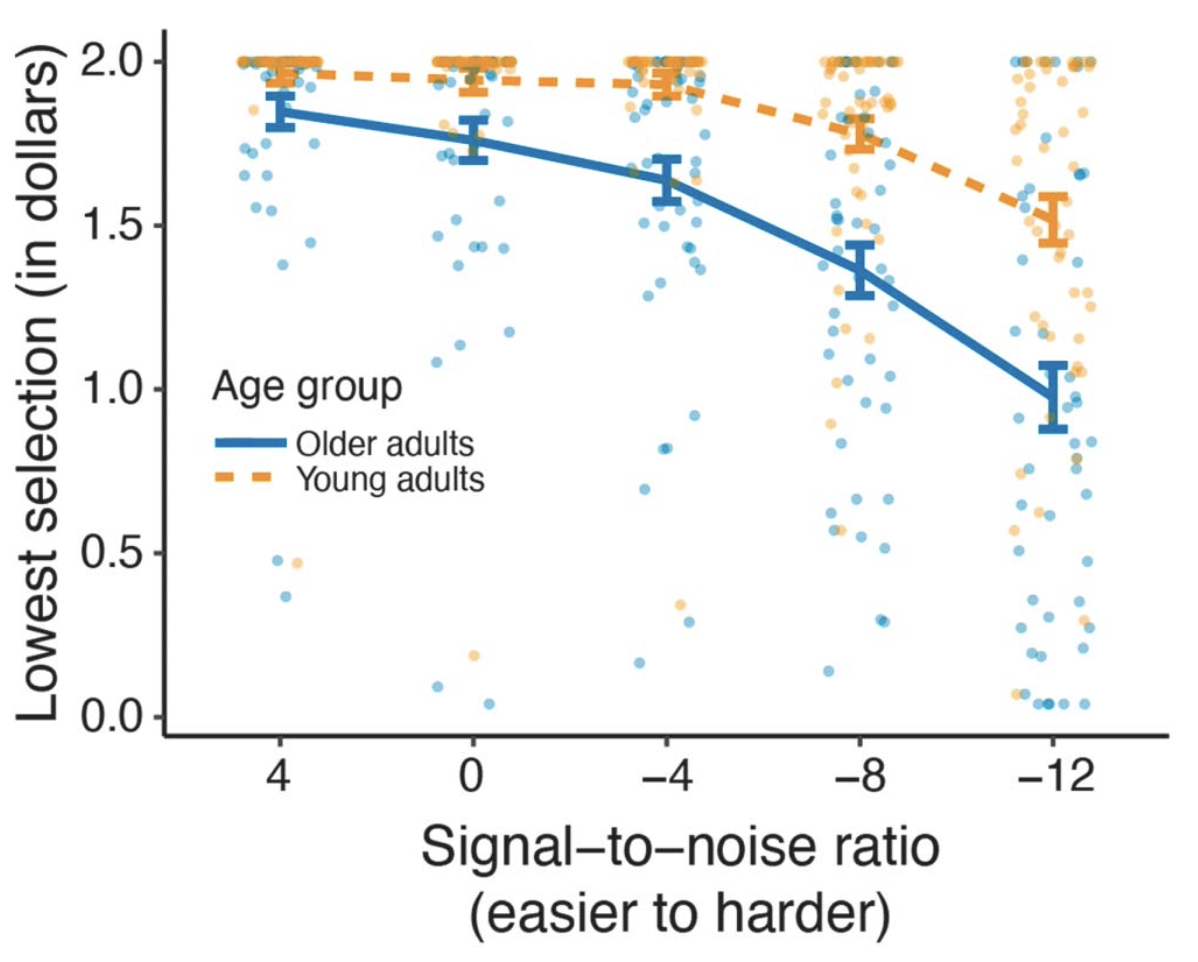
A common way to assess hearing sensitivity is the pure-tone audiogram—asking someone to raise their hand when they hear a beep, and figuring out the softest sound that can be reliably heard. Although this is useful information, it fails to capture the complexities of communication in real-life situations. Importantly, two people can have identical audiograms, but very different experiences: one could have no complaints about communication, whereas the other might frequently complain about not being able to have conversations in background noise. These self-report measures are an important component of hearing health care.
For the last few decades, self-reported communication difficulties have been known by the outdated term “hearing handicap”. Although hearing handicap is still found in the literature, I will use “auditory communication difficulties” (ACD) instead—it’s a mouthful but I think more accurate.
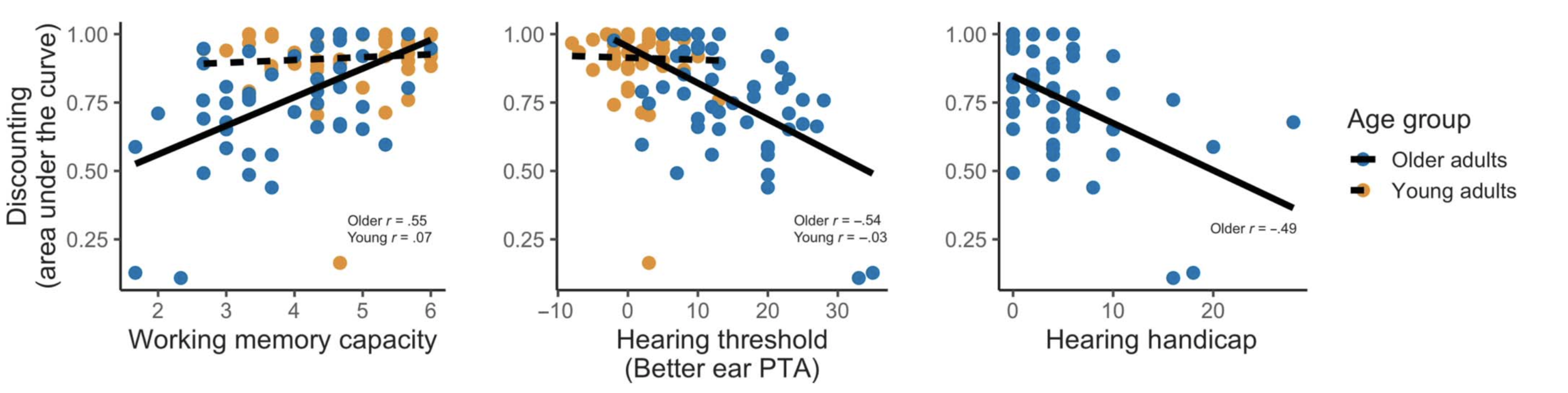
One area that has received surprisingly little attention is how auditory communication difficulties interact with mental health. Anxiety in particular seems relevant—people with hearing difficulty may experience greater anxiety in social situations than those without, or people with more baseline anxiety may feel more impacted by their hearing loss. To study this issue, we conducted a survey in which we asked people about their mental health (general anxiety, social anxiety, etc.) and auditory communication difficulties.
Characterization of outcome measurements and their relationship to each other. The diagonal shows score distributions for each outcome measure. Off-diagonal panels show either a scatterplot or Pearson correlation value for the variables indicated. *** = p < .001.
We found strong correlations between all of these factors. Of most interest was the correlation between auditory communication difficulties and social anxiety, which has not been shown before. We think this opens up a whole host of new possibilities for understanding everyday communication, and also treating communication challenges more holistically than have typically been done.
McClannahan KS, McConkey S, Levitan J, Rodebaugh TL, Peelle JE (2005) Social anxiety, negative affect, and hearing difficulties in adults. Trends in Hearing 29. doi:10.1177/23312165251317925